Where Is The Gender In Behavior Genetics? The Need For Social Epidemiology In Research On Gene-Environment Interactions
In cases where social environments are reflective of gender difference or inequality, social epidemiology in research on gene-environment interactions becomes more needed and more relevant.
Author:Suleman ShahReviewer:Han JuDec 18, 202315.4K Shares205.3K Views

How important is social epidemiology in research on gene-environment interactions?
Despite evidence of pervasive gender differences in morbidity and mortality, as well as gender-specific genetic association in some diseases, research on candidate gene-environment interaction is rarely informed by social scienceperspectives on gender and health.
Omitting basic theories of gender stratification from the study of social-environmental moderation of genetics may contribute to problems with replication and false-positive results in G×E research.
This article discusses the need for theories of gender and social inequality in research on gene-environment interactions in behavior genetics.
Preliminary Discussion
In the past decade, proliferation of genetic data has provided opportunities to examine interactions between genotypes and environments.
Yet, despite the promise of a genomics revolution, G×E research has fallen short of expectations for new epidemiological insights and targeted treatments.
Moreover, findings have proven difficult to replicate, and some argue that even independent replication may not provide sufficient evidence for true G×E effects.
In a recent critical review, evidence of strong publication bias towards positive findings combined with samples that are largely underpowered to detect small interaction effects led the authors to argue that “most, if not all, positive cG×E (candidate gene-environment interaction) findings represent type I errors (1041).”
These authors conclude that enthusiasm about novel G×E findings has been misplaced.
Despite on-going skepticism, many researchers remain optimistic about the promise of an integrative paradigm that sees G×E as a fundamental process in disease pathways.
Failure to replicate genetic associations, including G×E effects, does not necessarily indicate that findings are an artifact of chance.
Rather, the replication problem may reflect a compound genetic architecture that is further obscured by an even more complex environment.
Population differences in environmental effects or allele frequencies, as well as interactions between multiple genes and environments, can render even strong general G×E effects nearly impossible to detect in independent samples.
As a G×E effect is unlikely to operate uniformly across all genetic and environmental contexts, a limitation of existing research is overreliance on simple models that do not accurately reflect the complexity of gene-environment interplay.
Consistent with this reasoning, G×E research rarely draws from advances in social science on the role of social forces in health and illness.
This failure to integrate and adjust methodologically to basic theories of social stratification in the study of interactions between genetic risk and socially-structured experiences and environments may contribute to problems with replication and false-positive results in G×E research.
Some examples of socially-structured experiences/environments include:
- family context
- parenting
- social support
- stress
- traumatic lifeevents
Recent doubt regarding our ability to reliably detect and report G×E poses challenges to the translation of G×E research, delaying potentially groundbreaking advances in personalized treatment and prevention.
In this critical review, I propose a framework for investigating more complex social pathways in G×E, arguing that moderation of biological processes by social environments is systematically moderated by gender.
Main Discussion
A framework for studying gender-moderated G×E effects (i.e., G×E×Gender) is advocated.
As G×E may be conditioned on gender or other social statuses associated with systematic inequalities in risks and resources, modeling interactions between genotype and one proximal indicator of the social environment is overly simplistic.
Gender can moderate G×E through at least three pathways:
(1) by stratifying men and women into different environments
(2) by differentially shaping the experiences of men and women in similar environments
(3) by influencing distinct biological, psychological, or behavioral responses to similar experiences
The importance of endophenotypes in identifying the timing and sequencing of gender moderation is discussed, and methodological considerations are offered to guide future research.
Why Gender?
There has been very little discussion of gender in G×E research.
This continues despite conclusions about the most promising candidate genes for major diseases often being specific to men or women, and evidence of substantial and enduring gender differences in morbidity and mortality.
The term “sex,” used frequently in G×E research, has historically referred to biological and physical characteristics that have a genetic basis in sex chromosomes.
“Gender,” however, is a social construction encompassing cultural conventions, roles and behaviors adopted by men and women that shape their experiences and activities.
Men’s and women’s opportunities and decisions are in part constrained by:
- institutions
- norms
- policies
- social structures
Over time, these constraints lead to gender differences in health and behavior that create, sustain, or intensify underlying biological sexual dimorphisms.
In practice, sex and gender often overlap in meaningful ways, such that differences between men and women are attributable to a combination of social and biological forces that are often difficult to disentangle.
Consequently, sex/gender is a theoretical dichotomy that delineates and reifies disciplinary boundaries, but is rarely defensible in application to illness and disease.
Here, I employ the term “gender” and focus in large part on social epidemiology, leaving discussions of straightforward biological sexual dimorphism to other reviews.
This choice is not intended to deny or subjugate the role of biology, but highlights the cumulative effects on men and women of living in a social world, where such categories matter.
Gender inequalitypersists in industrialized countries despite economic, political, and ideological changes that might have leveled the playing field for men and women.
Because people use gender as a primary frame for organizing social relations, and therefore for shaping their own behavior and sense of self, gender inequality is continually incorporated into emerging social structures in ways that influence health.
Gender, therefore, is an enduring fundamental social cause of disease.
Health consequences of gender stratification are reflected in:
a. Social roles
For example, reduced physical activity associated with motherhood, but not fatherhood.
b. Social relationships
For example, stress associated with the unequal division of household labor among married couples.
c. Social policies
For example, inadequate parental leave that disrupts women’s career trajectories and lifetime earnings.
They differentially structure men’s and women’s access to health-promoting resources and exposure to risk factors.
Gender shapes virtually all health outcomes, working through innumerable more proximal pathways that may be, in nature:
- biological
- psychological
- behavioral
A Framework For Gender-Inclusive Research In Behavior Genetics
Gender Moderation Of G×E
While men’s and women’s autosomal genomes are very similar, there are marked gender differences in gene regulation and expression.
Although seldom modelled to reflect such complexities, G×E effects may be conditioned on gender or other social statuses associated with systematic inequalities in risks and resources.
A statistical interaction is only identifiable when there is sufficient variation in genotype and environment across the distribution of relevant social categories, suggesting that modeling interactions between genotype and one proximal indicator of the social environment is overly simplistic.
Yet, social environmental effects are almost always assumed to be uniform across gender groups in G×E research, even when the social science literature points to gender differences in social epidemiology.
This is highly problematic because differences in the susceptibility and consequences of social experiences for men and women can lead to false-negative results in G×E models, particularly in smaller samples typical of replication studies.
In cases where social environments are reflective of gender difference or inequality, hypotheses on G×E must be gender-specific (i.e., G×E×Gender), presenting both unique challenges and enormous potential for social epidemiology in transdisciplinary agendas on health and behavior.
Gender can influence environmental moderation of genotype through at least three distinct mechanisms.
First, men and women may be systematically subjected to distinct environmental risk and protective factors.
For example, men and women are exposed to different levels and types of environmental toxins due to gender differences in occupations and household division of labor.
Also, there are gender disparities in exposure to chronic strain and traumatic life events that interact with genetic risk factors in disease pathways.
Women are disproportionately susceptible to socioeconomic stressors (i.e., poverty and joblessness), as well as exposure to sexual and intimate partner violence.
In contrast, men are substantially more likely than women to be victims of all other kinds of violent crime, and are more susceptible to stressors associated with the workplace and the military.
There are important gender differences in access to protective resources as well.
For example, religiosity and social support activation are stronger among women relative to men.
These kinds of patterns may result in insufficient variation on certain environmental risk and protective factors for either men or women, making it difficult to identify a G×E interaction without introducing gender moderation into the model.
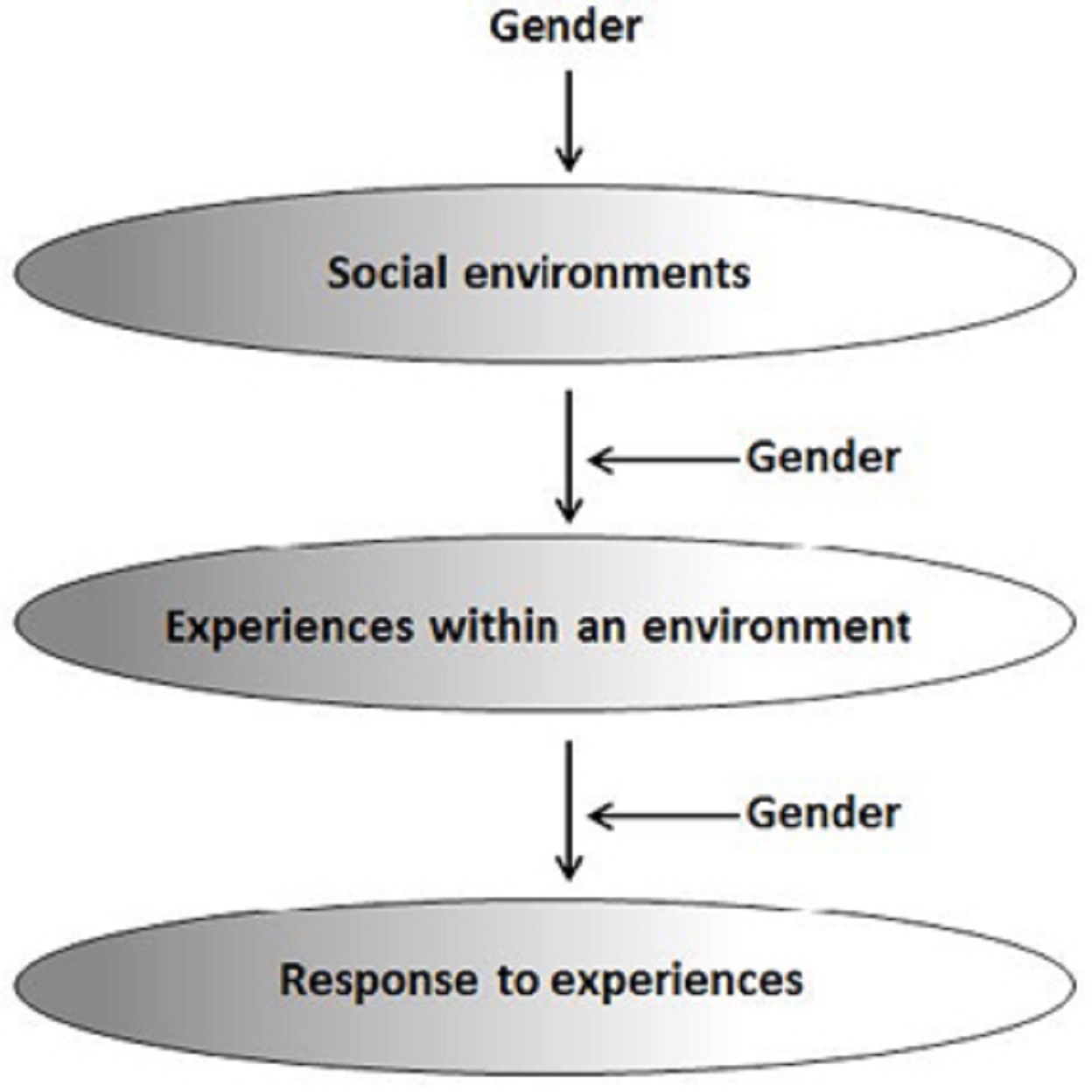
Second, men and women may have qualitatively different experiences even when embedded in the same environments.
Because G×E research often employs simple measures of the presence or absence of environmental risk and protective factors, distinctions between men’s and women’s experiences that could differentially shape health outcomes are masked.
For example, compared to being childless, being a custodial parent has been linked to higher psychological distress among women, but lower levels of substance use and abuse among men.
These trends may be explained by gender differences in the experience of parenthood.
The vast majority of people who experience child-rearing as single parents are women, and this status is strongly linked to:
- poverty
- unemployment
- other adversities
Even within married couples, most mothers are the primary caregivers to their children. For that, when compared to fathers, mothers engage in substantially more:
- multitasking
- physical labor
- emotional parenting work
In other words, while both men and women reap psychosocial rewards from parenting, the burdens are disproportionately shouldered by women, potentially leading to gendered health-promoting or protecting functions.
In cases like this, environments are likely to have unique interactions with genotypes for men and women because they are experienced in fundamentally different ways, resulting in a G×E that operates exclusively or more strongly in one gender group.
Third, men and women may have different responses to the same experiences.
Gendered responses to similar experiences may be biological, psychological, or behavioral in nature.
For example, research suggests that women are significantly more reactive to stress compared to men, as measured by both physiological and subjective responses, particularly if stressors involve relationships.
In addition, there is some evidence that men are more resilient to environmental stressors than women because they are socialized to capitalize on heritable psychosocial resources like:
- active coping
- mastery
- self-esteem
Conversely, women exhibit a stronger propensity to seek both informal social support and health services in response to stress.
These gender differences in reactivity likely moderate genetically-influenced pathways linking stressful life events to physical and mental illness.
Thus, even when men and women occupy the same environments and have similar experiences, they bring gendered expectations, identities and resources that differentially affect their health-related reactions.
Role Of Endophenotypes
Understanding the timing and sequencing of gender moderation in specific epidemiological pathways is critical for effective clinical translation and policy development.
Incorporating endophenotypes (i.e., intermediary biomarkers that link genetic variability to complex phenotypes such as psychiatric disorders) provides insight into when and how gender shapes disease pathways.
Specifically, gender may influence moderating environments before endophenotypic expression through regulation of gene expression as synthesized proteins.
Environmental factors can cause hormonal or neurological changes that alter gene transcription or translation. Such factors include:
- chronic stress
- poor nutrition
- exposure to toxins
In cases where gender moderation occurs prior to endophenotypic expression, men and women with the same genetic and environmental profiles will have different endophenotypic outcomes.
As the presence of the endophenotype will depend on both gender and environment, patterns will reflect a stronger or exclusive effect of genotype on endophenotype and phenotype in a particular environment for either men or women.
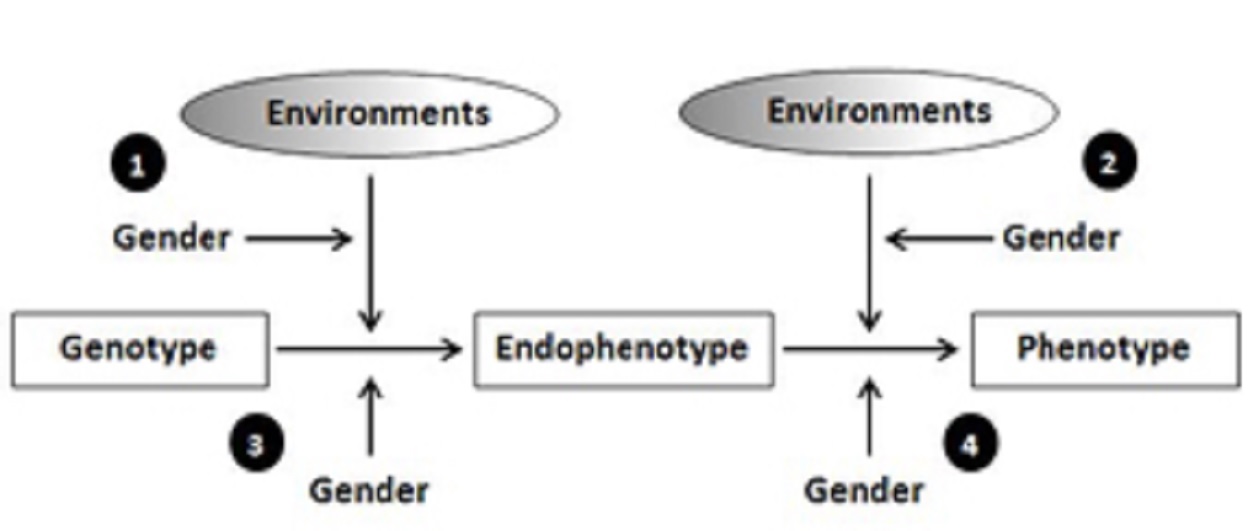
In contrast, gender can also moderate genetic risk or G×E through purely social mechanisms after endophenotypic expression, reflecting:
- social inequality
- social control
- other normative influences
When this occurs, men and women with common genetic risk factors will display similar endophenotypes.
However, social influences that differ systematically by gender in their intensity or effect will lead to unique phenotypic outcomes for men and women.
This pathway may result in a pattern in which men or women are more likely to demonstrate a particular phenotype or set of phenotypic outcomes in a given environmental context (i.e., no effect or reduced effect for one gender).
However, equally plausible is that the same endophenotype may lead to unique, gendered phenotypes in response to similar environmental risk and protective factors.
In addition to pathways involving social environmental influence, gender can moderate genetic risk directly, independent of environments.
Interactions between gender and genotype may be due to hormonal or other biological differences in gene regulation or phenotypic expression.
In this case, a G×E may be observed for only one gender group, but this is an artifact of sexual dimorphism through gene transcription or translation prior to an endophenotype or an unmitigated effect of gender after development of an endophenotype and prior to the emergence of a phenotype.
In other words, rather than gender working with or through different environments, a G×E may be specific to men or women only because a genetic factor does not confer risk for one group under any environmental conditions.
Empirical Illustration
To demonstrate how G×E×Gender pathways might function in disease outcomes, I draw from research on the social and genetic epidemiology of substance use.
In this case, gender moderation is thought to emerge after endophenotypic expression, leading to different phenotypic outcomes in men and women.
Specifically, the high-risk variant of GABRA2 (gamma-aminobutyric acid A receptor, alpha 2) is believed to increase risk for an impulsivity endophenotype associated with compulsive maladaptive behavior in response to heightened emotional states.
In environments characterized by high levels of social regulation (e.g., religious upbringing, high parental monitoring), individuals are less likely to engage in alcohol use irrespective of endophenotype or genotype on GABRA2.
Conversely, when people are embedded in more permissive or negative social groups, patterns of drug and alcohol use reflect the full range of genetic and endophenotypic variation (i.e., G×E).
However, recent research suggests that these G×E patterns in alcohol dependence operate only among men, constituting a G×E×Gender effect.
Furthermore, work in progress suggests that this gender moderation also emerges in smoking and drug dependence outcomes.
Findings point to gender-specific suppression of a GABRA2 effect on substance use through stigma and gender prescriptions that discourage certain kinds of problematic drinking and drug use among women and mothers.
In this case, gender moderation of the gene-environment interaction likely occurs after endophenotypic expression.
Consistent with this pattern, recent research finds that the high-risk variant of GABRA2 and the associated impulsivity endophenotype increase risk for obesity among women, but not men.
While interactions between GABRA2 and the social environment in obesity pathways have yet to be tested, it is plausible that gender norms influence men and women with the same genetic and environmental risk factors to engage in different maladaptive behaviors (i.e., overeating versus substance abuse).
The case of GABRA2 and social regulation constitutes early empirical evidence for the feasibility and importance of research on G×E×Gender.
It also demonstrates that more complex theory and analyses are needed to identify better estimates of both genetic and environmental effects.
In analyses predicting alcohol dependence, the straightforward G×E is entirely masked by gender differences in the direction and magnitude of the environmental effect.
Moreover, in the case of smoking, no main effect of genetic risk on GABRA2 is identified in the model without the gender interaction.
Yet, when gender moderation is introduced, the effect of GABRA2 and its interaction with the environment are robust and strong in magnitude.
Although more research needs to be done in this area, these relationships are likely to be the norm rather than the exception given the pervasive influence of gender in social epidemiology.
Considerations For Future Research
Going forward, overcoming doubts about the reliability and feasibility of G×E will require serious engagement with social theories of status and inequality.
To date, social categories that stratify individuals into different environments and shape their experiences within those environments have been virtually ignored.
The theoretical framework laid out here can serve as a starting point for future theory development and empirical research on gender moderation of G×E effects in complex, heritable disorders.
In a study published in the journal JAMA Psychiatry(when it was still called Archives Of General Psychiatry) in 2005, its authors concluded that it is important to highlight considerations for minimizing bias and errors in future research on G×E×Gender.
Prior to data analysis, a comprehensive review of the behavior genetics and social science literatures should be conducted to identify and hypothesize plausible G×E×Gender pathways a priori.
In addition, the following variables should be carefully selected or constructed to reduce measurement error and increase power to detect significant relationships:
- endophenotype
- phenotype
- environmental variables
Moreover, the adequacy of variation across combinations of these variables should be assessed.
Furthermore, the presence of rGE and possibility of gender-moderated rGE (i.e., genetically-influenced environments that produce distinct phenotypic outcomes in men and women) should be examined.
As three-way interactions are more complicated to evaluate statistically than traditional two-way G×E, a careful analysis plan is critical.
The following should be tested with a series of three stepwise regression models using a pooled sample:
- main effects (Gender, G and E)
- two-way interaction effects (E×Gender, G×Gender, G×E)
- three-way interaction effects (G×E×Gender)
For two-way and three-way interactions, Chow-type tests of the equality of coefficients across groups are required to determine statistical significance at p < 0.05.
When using logit models for dichotomous outcomes, Chow tests confound the magnitude of the effect for each group with group differences in residual variation.
Consequently, differences in predicted probabilities should be examined using Long’s delta method.
In addition, figures of predicted values or probabilities of phenotypic expression at different levels of genotype and environment must be presented to demonstrate effect size.
Adjustment for multiple testing should be considered if the sample is large enough to balance concerns about Type II errors.
Together, these strategies should provide a robust and conservative test of G×E×Gender.
Following an analysis of G×E×Gender, extension of effects beyond the original environment, candidate gene, endophenotype and phenotype combination should be evaluated.
If a significant three-way effect is found, this strategy will help identify polygeny or pleiotropy.
Additionally, replication should be conducted using one or more independent datasets to alleviate concerns about publication bias and false-positive results.
Conclusion
In sum, the proposed framework promotes a shift in genetic epidemiology towards research that accounts for distal social and proximate biological processes in equal measure, emphasizing the power of gender to moderate environmental and genetic influences.
For decades, the distinction between sex and gender has served to delineate the domains of biology and social science, creating an intellectual space for the study of hormonal and chromosomal sexual dimorphism on one hand, and socially constructed difference and inequality on the other.
However, relegating biological differences between men and women to sex, as something distinct from gender, has undersold the role of culture and social inequality in producing those differences.
Often, gender structures men’s and women’s health and wellbeing through social as well as biological pathways.
That is, biological characteristics and processes interact with gendered social roles and relations, which shape patterns of exposure to stress and other risk factors, differential access to resources for improving or maintaining health, and the unique meanings that various structural positions and experiences have for men and women.
Therefore, social epidemiology in research on gene-environment interactions must be considered.

Suleman Shah
Author
Suleman Shah is a researcher and freelance writer. As a researcher, he has worked with MNS University of Agriculture, Multan (Pakistan) and Texas A & M University (USA). He regularly writes science articles and blogs for science news website immersse.com and open access publishers OA Publishing London and Scientific Times. He loves to keep himself updated on scientific developments and convert these developments into everyday language to update the readers about the developments in the scientific era. His primary research focus is Plant sciences, and he contributed to this field by publishing his research in scientific journals and presenting his work at many Conferences.
Shah graduated from the University of Agriculture Faisalabad (Pakistan) and started his professional carrier with Jaffer Agro Services and later with the Agriculture Department of the Government of Pakistan. His research interest compelled and attracted him to proceed with his carrier in Plant sciences research. So, he started his Ph.D. in Soil Science at MNS University of Agriculture Multan (Pakistan). Later, he started working as a visiting scholar with Texas A&M University (USA).
Shah’s experience with big Open Excess publishers like Springers, Frontiers, MDPI, etc., testified to his belief in Open Access as a barrier-removing mechanism between researchers and the readers of their research. Shah believes that Open Access is revolutionizing the publication process and benefitting research in all fields.

Han Ju
Reviewer
Hello! I'm Han Ju, the heart behind World Wide Journals. My life is a unique tapestry woven from the threads of news, spirituality, and science, enriched by melodies from my guitar. Raised amidst tales of the ancient and the arcane, I developed a keen eye for the stories that truly matter. Through my work, I seek to bridge the seen with the unseen, marrying the rigor of science with the depth of spirituality.
Each article at World Wide Journals is a piece of this ongoing quest, blending analysis with personal reflection. Whether exploring quantum frontiers or strumming chords under the stars, my aim is to inspire and provoke thought, inviting you into a world where every discovery is a note in the grand symphony of existence.
Welcome aboard this journey of insight and exploration, where curiosity leads and music guides.
Latest Articles
Popular Articles